您好,欢迎来到显微镜配件耗材官网!
 最新资讯返回
最新资讯返回人脑可以说是宇宙中最复杂的结构之一。它由超过1000亿个神经元组成,可以执行高度复杂的计算。重要的是,它能够不断适应新的情况,让我们形成对我们经验的回忆。在大脑中,神经元彼此连接,每个神经元有多达10,000个突触。突触联系的形成和消除以及现有突触的功效可以通过经验来修改。这个动态的过程被称为突触可塑性,对学习和记忆等高级脑功能至关重要。了解突触传递和可塑性的分子机制是神经科学的基本问题之一。
突触传递是由从激活的突触前神经元向突触后受体释放的神经递质的结合介导的。 GABA和甘氨酸是与抑制性受体结合的主要抑制性神经递质,谷氨酸是哺乳动物脑中主要的兴奋性神经递质。尽管代谢型谷氨酸受体(mGluRs)和NMDA受体(NMDARs)发挥更大的调节作用,AMPA受体(AMPARs)是中枢神经系统中快速兴奋性突触传递的主力。 AMPARs是由四个亚基GluA1-4的不同组合组成的四聚体配体门控离子通道。
突触可塑性被广泛认为是学习和记忆的细胞基础。它涉及神经元的分子和细胞结构的长期变化,其决定了如何容易地产生响应于特定突触前输入的动作电位。突触可塑性研究最多的形式是长时程增强(LTP)和长时程抑制(LTD),其中突触传递的强度分别增加和减少。在分子水平上,LTP和LTD都是由突触后膜中AMPARs的数量和活性的持续变化介导的。因此,调节AMPAR膜运输是突触可塑性,学习和记忆的关键细胞机制[1,2]。
AMPAR进出突触是一个动态的过程,涉及内质体运输以及沿质膜横向扩散[3]。中断适当的AMPAR功能或贩运损害突触可塑性,因此,学习和记忆。 AMPAR或调节AMPAR运输的蛋白质的遗传改变可能与多种疾病相关,包括自闭症谱系障碍,精神分裂症,阿尔茨海默病和智力障碍[4]。因此,理解AMPAR运输中依赖经验的变化的分子过程对于理解行为的细胞基础以及许多神经疾病是至关重要的。在这篇综述中,我们将讨论AMPAR运输领域的重要发现,重点研究在原代分离的培养神经元或器官型脑切片培养物(体外)和活体动物(体内)中使用延时和纵向成像方法的研究。
生物化学,细胞生物学和电生理学工具已广泛用于研究突触可塑性,并大大提高了我们对潜在细胞和分子机制的认识。但是,光学显微镜可以直接观察细胞和分子。在固定的脑片和解离的神经元培养物中的免疫组织化学已经揭示了AMPAR在基础和刺激条件下的空间分布[5-7]。但是,由于AMPAR运输的动态性,只有实时成像可以实时监测个体突触的变化。
允许AMPAR蛋白延时成像的重要技术进展是用荧光蛋白标记AMPAR亚基(图1a)。在确认绿色荧光蛋白(GFP)与AMPAR融合不影响其通道特性后,Shi等人[8]将GFP标记的AMPARs在细胞培养和脑切片中成像,并显示表达模式模拟内源蛋白的分布。他们使用这种方法,观察到在基础状态下含有少量GluA1亚基的受体在树突中,但是在强直性突触刺激下,GluA1在树突中并沿着树突棘聚集。
GFP标记的AMPAR虽然有用,但不能用于区分细胞内受体和插入质膜的受体, 而这种区别至关重要,因为只有膜结合的AMPARs是活跃的并且参与突触传递。为了选择性地观察膜插入的AMPAR,Ashby等人[9••]使用对酸碱度敏感的pHluorin(SEP)而不是GFP来标记AMPARs。 SEP是GFP 的 pH敏感型,在中性pH值(7.4左右)下荧光较亮,但在酸性环境中淬灭[10]。 AMPAR插入和脱出细胞质膜是由胞吐和胞吞介导的[11,12]。在细胞内,AMPARs穿过分泌和内吞途径的酸性环境。当存在于膜上时,它们暴露于pH中性细胞外空间。因此,通过将SEP插入到AMPARs的细胞外或内腔暴露的末端(N-末端),大部分荧光信号将源自细胞表面的受体(图1a和c),其中一小部分来自AMPARs的荧光信号内质网的pH中性内腔[13]。
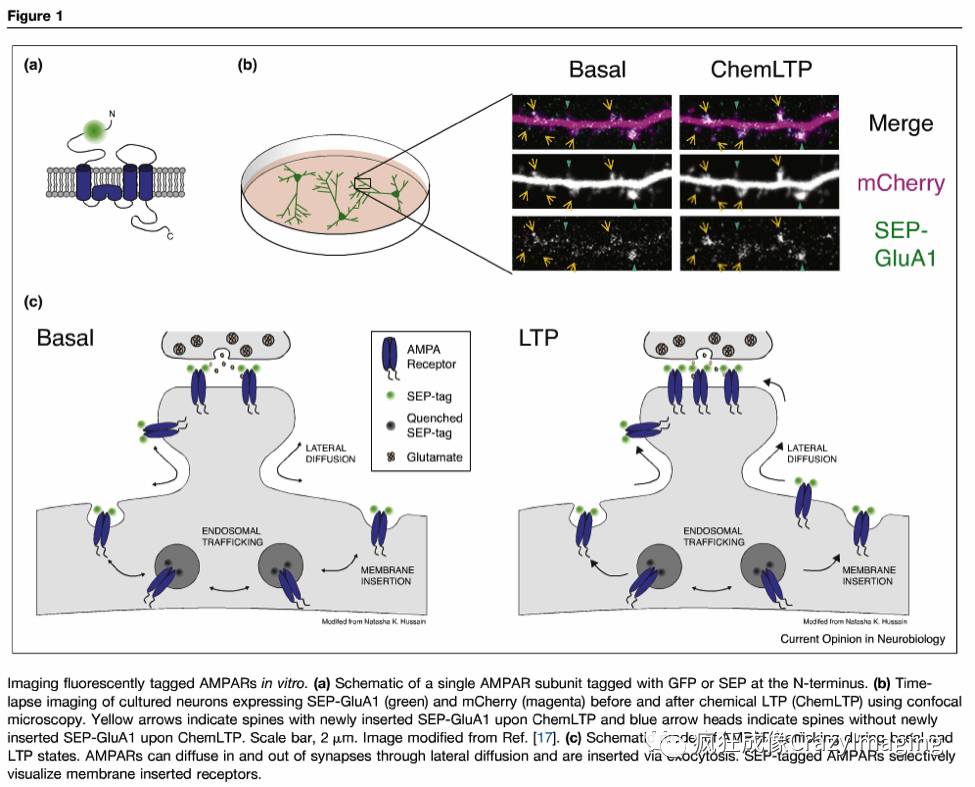
这种方法允许研究不同的AMPAR亚基的表面定位,LTP-或LTD-诱导刺激后AMPAR的膜运输,以及离散插入事件的可视化。在分离的神经元培养物和海马切片培养物中,通过应用甘氨酸或 Forskolin(化学LTP)的孵育,或者通过在单一的棘突附近释放谷氨酸,可以药理学诱导突触增强。这些系统已被用来表明在LTP诱导后GluA1和GluA2的表面表达增加[14-17](图1b和c)。在棘突表面添加AMPARs可以通过树突轴的横向扩散或通过细胞内隔室的胞吐作用发生(图1c)。区分这两种机制的一种方法是将SEP标记的AMPAR的延时成像与光漂白后的荧光恢复(FRAP)相结合。在大范围内对AMPAR表面群体进行光漂白后,只有新胞吐的受体才可见[18,19]。尽管横向扩散不断补充AMPAR在棘突[19,20,20,21],LTP后表面AMPAR的增加需要胞吐[15]。然而,AMPARs的胞吐作用是直接发生在棘突还是邻近棘突的树突上,然后是侧向扩散到棘突中仍然存在争议[3]。使用全内反射荧光(TIRF)显微镜,可以直接观察AMPARs在原代神经元培养物中的单个插入事件[22,23,24]。这些研究表明大多数插入事件发生在体细胞和树突状细胞中,并且从这些插入位点释放的AMPARs可以扩散到棘突中。然而,在突触活动或LTP期间,直接在棘突中也可观察到胞吐[14,25,26]。
NMDAR激动剂NMDA在培养的神经元中的应用诱导了突触的长时程抑制(化学 LTD)[27]。在这个过程中成像SEP标记的AMPARs已经显示出表面受体的快速细胞减少,然后缓慢恢复[9]。 LTP和LTD实验已经被用于研究蛋白质 - 蛋白质相互作用和翻译后修饰在AMPAR运输中的作用[28-31]。然而,使用NMDA诱导的回收实验需要仔细控制,因为应用NMDA刺激也会诱导细胞内区室的快速酸化,从而使来自SEP标记的AMPARs的荧光强度变化的解释变得复杂化[32]。使用荧光标签进行成像受体运输的另一个问题是,这通常涉及过度表达单个AMPAR亚基并且具有与受体蛋白质的N末端融合的荧光蛋白。这可能影响突触受体的亚基组成,荧光标记可能干扰AMPAR亚基之间的相互作用,或者AMPAR与辅助亚基如跨膜AMPAR调节蛋白(TARPs)家族之间的相互作用。避免标记受体过度表达的解决方案是产生敲入小鼠或神经元,将其表达的内源性受体用荧光蛋白标记。
另一种可视化AMPAR在神经元膜中的横向运动并避免使用过度表达荧光蛋白融合AMPAR的方法是单粒子追踪。用荧光染料或量子颗粒(quantum dots, QDs)标记针对GluA1或GluA2的抗体,并稀疏地施加到培养的神经元的表面。 AMPAR运动的实时跟踪揭示了单个含GluA2的AMPARs要么静止要么快速扩散[32]。与突触外受体相比,突触内受体表现出更多的扩散受限[33]。在突触活动期间,含有GluA1的AMPAR在降低运动性后被捕获并积聚在突触位点[34],并且脱敏的AMPAR与带正常功能的受体的横向交换可以使突触抑制后的突触传递得到恢复[20]。这些使用QD追踪AMPAR运动性的研究深化了我们对单个受体蛋白质行为的了解,然而,使用这种方法的主要缺点是使用QDs通过抗体标记受体使受体增加了体积较大的复合物(约50nm),而这可能干扰其在空间狭窄区域的转运,例如宽度约30nm的突触间隙[35-38]。事实上,研究表明,通过使用小荧光染料[35],减小量子颗粒尺寸[36,37],或通过使用生物素和链霉亲和素直接连接量子点[38],观察到的受体动力学也会发生改变。
AMPARs在突触的限制性扩散可以通过在解离的神经元培养物中,用SEP标记的AMPARs在单个部分光漂白之后进行成像来进一步表现 [39]。此外,最近已经使用超分辨率显微镜成像显示突触后密度(PSD)是由突触后支架蛋白如PDS-95簇形成的纳米级亚结构域中构建的, 而AMPARs集中在可能有助于调节突触传递的这些簇中。这些研究暗示,不仅AMPARs突触内外交换,而且在突触内移动也可能会在突触传递和可塑性中发挥重要作用[40]。
除了更好地了解AMPAR运输的动态之外,活体成像还提供了研究涉及调节AMPAR运输的分子机制的方式。通过在AMPAR本身或相互作用的蛋白质上引入遗传缺失或突变,许多研究已经证明磷酸化对AMPARC-末端以及架构和运输蛋白的适当结合的重要性,以及辅助性亚基例如TARP对于适当的AMPAR运输[16,22,28]。有关AMPAR运输机制的进一步综述,请参见[2,41,42]。
如上所述,在解离的培养的神经元和切片培养物中已经实现了AMPAR的成像。然而,这些制剂的离体性限制了这些发现的解释。为了在获取和维护学习任务期间随着时间的推移追踪个体神经元和个体棘中AMPAR水平的变化,AMPAR的可视化需要移动到完整的动物中。
成像方法,特别是双光子激光扫描显微镜(TPLSM),近年来被广泛用于研究突触可塑性。 TPLSM是在25年前发明的[43],能够在完整的神经组织中实现高分辨率和高灵敏度的荧光成像。在过去的20年中,TPLSM已经被用于在细胞和亚细胞水平上观察活体动物中的神经元结构以及神经元活动。许多研究使用基因编码的荧光钙指标如GCaMP系列来显示神经元活性[44]。这些研究对我们对小鼠行为期间功能性神经元网络及其活性的理解作出了重大贡献。
此外,使用TPLSM和神经元中表达荧光形态学标记的转基因小鼠,研究人员在发育和学习期间研究了几天到几个月的结构变化[45]。大量的研究集中在树突棘,突触后兴奋性突触所在的部位。虽然从固定样品的研究中已知树突棘密度会随着时间的推移而改变,但是现在可以通过重复成像相同的神经元并追踪个体树突棘随着时间的变化来直接研究树突棘更新率[45-50,51] 。这些研究发现,树突棘在体内确实是动态的。它们在数分钟,数小时或数天内出现和消失,并在最早的可塑性时期之后在动物成体中变得更加稳定。此外,感官剥夺或富集以及学习可以改变树突棘周转率。
体内结构成像已经提高了我们对树突棘周转和树突重塑的理解,这可能是在发育和体验中重新生成神经连接的基础。然而,为了解决突触可塑性的详细机制,如LTP或LTD,我们需要转向分子。几个小组通过用荧光蛋白例如GFP标记感兴趣的蛋白质在体内可视化分子。 Gray等人研究了年轻小鼠的突触后支架蛋白PSD-95,发现在早期发育过程中,树突棘中的PSD-95簇可以保持稳定[52]。然而,使用标记有可光激活形式的GFP的PSD-95,它们显示突触PSD-95被迅速重新分配,并且PDS-95的突触滞留时间随着发育年龄增加并且在感觉剥夺后减少。进一步的研究表明,随着时间的推移,树突棘中的胞浆树突棘体积和PSD-95-eGFP水平会随着时间的推移而变化,这表明许多个体突触的强度在成年新皮层中不断变化[52,53]。
成像分子还允许研究缺乏明显形态结构的抑制性突触,如作用于兴奋性突触的树突棘。使用荧光标记的Gephyrin(抑制性突触处的结构蛋白)研究表明,树突状突起和棘突上的抑制性突触在树突状分叉中的分布以及在正常和改变的感官体验期间的重塑动力学方面不同[54,55]。此外,他们发现抑制性突触和树突棘重塑在空间上聚集,并且受感官输入的影响[56]。
虽然可视化树突棘结构和突触结构蛋白在体内已经揭示了重组突触输入到神经元的动态事件,关于功能相关性,分子组成和突触强度的信息仍然是难以捉摸的。为了更好地理解经验和行为如何塑造突触强度,我们需要进行突触受体的体内成像。
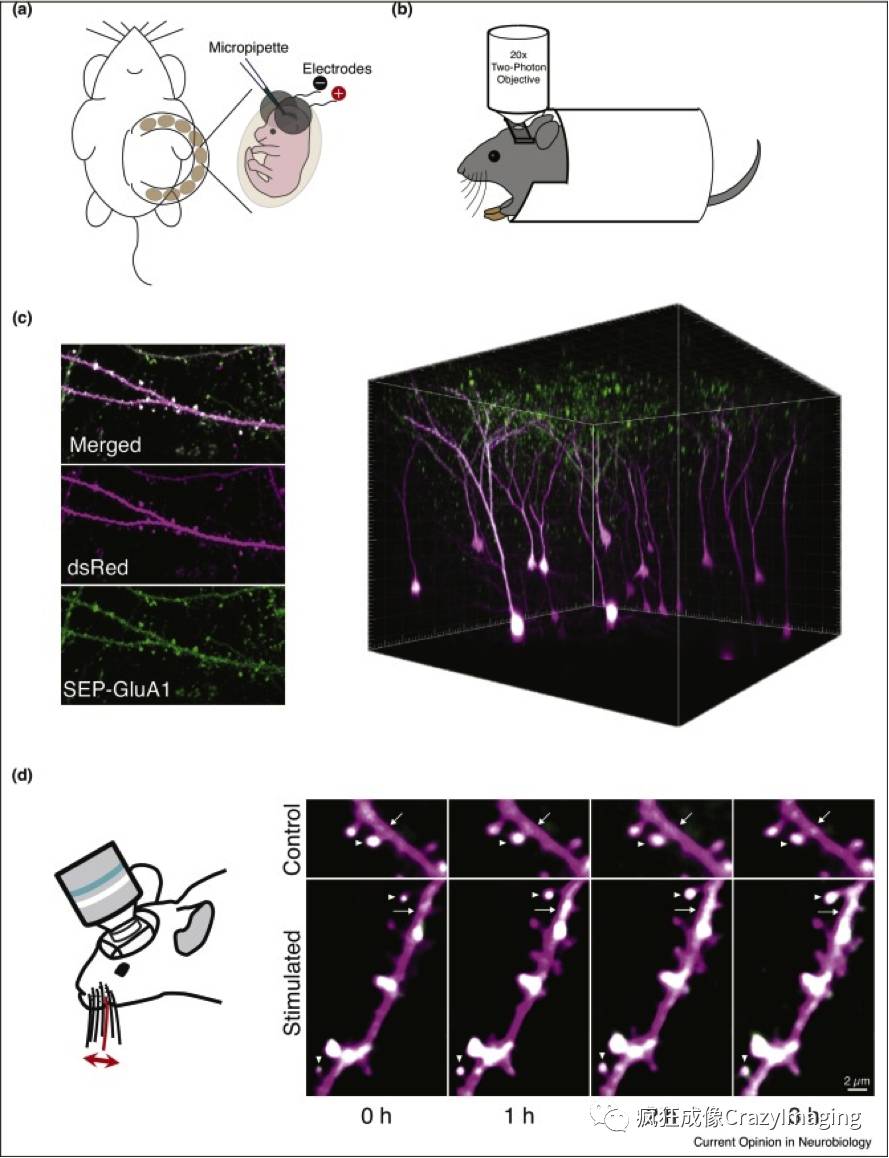

由于技术上的困难,AMPARs的体内成像直到最近还没有实现。通过在子宫内电穿孔(图2a)和双光子成像(图2b)相结合,我们小组能够在活体小鼠的躯体感觉皮层的2/3层神经元的突触中观察到SEP标记的GluA1(图2c)[57••]。虽然我们的研究使用基于Sutter MOM系统和ScanImage软件的显微镜进行[58],但目前大多数商用双光子显微镜系统能够在体内可视化荧光标记的受体。这种方法使得能够在基底期间以及在受刺激条件下纵向追踪神经元个别突触处的AMPAR。我们发现快速刺激小鼠胡须导致神经元表面AMPAR GluA1亚基(sGluA1)在树突棘和周围的树突状细胞中的强度显着增加,同时仅产生树突棘尺寸的小变化(图2d)。有趣的是,树突棘sGluA1强度的变化与胡须刺激后树突状细胞sGluA1强度的变化呈正相关但不同。胡须刺激诱发的棘突sGluA1强度增加持续时间长,且与NMDAR有关。以前,树突棘结构动力学已被用作突触可塑性的量度。然而,这些结果表明,通过单独研究树突棘的周转率,可能会忽略先前存在的树突棘的关键可塑性事件。除了研究树突棘动力学和树突棘尺寸变化以外,AMPAR成像还可以显示树突棘内发生的可塑性。这些数据表明,在先前存在的棘突的可塑性比树突棘的更新分布更广泛,成像研究在体受体的能力开辟了一个新的,更敏感的方式来研究突触可塑性。
下一步将是研究AMPAR动态在体内的小鼠接受学习任务。已经显示许多学习任务通过电生理记录在大脑的任务相关区域中诱导突触增强。通过在这些大脑区域的神经元中对AMPAR进行成像,我们将了解分子水平上的突触可塑性如何导致对细胞群体水平和最终行为的影响。
荧光标记的AMPARs被广泛用于在突触可塑性期间直接观察AMPAR在神经元中的动态。在体外的AMPAR运输成像研究已经使我们深入了解了AMPAR 在突触内外交换的调控机制,而将在体内AMPAR运输可视化揭示了受体在收到感官刺激时的动态变化。体外研究提供了方便于通过药理学或电刺激操纵的巨大优势,因此非常适合于进一步研究AMPAR运输和突触可塑性的详细机制。体内系统允许监测当动物正在经历或学习新的任务时突触的变化,并可以提供突触分子变化与行为之间的直接联系。此外,与离体制备的样品相比,整个脑网络在体内是完整的,并且神经元正在接收并整合各种各样的生理信号。将突触AMPARs的可视化与用于细胞内信号事件和钙成像的光学传感器成像相结合的研究将能够揭示神经元中的信号事件如何导致突触可塑性以及突触可塑性如何能够形成神经元的放电模式。体内的神经递质受体运输等分子事件也提供了一个有用的工具,可用来研究那些编码影响突触可塑性的蛋白质的基因突变如何影响大脑和行为如学习和记忆的变化。
 QQ 501747125,QQ 3318510886网站建设:合优 网易企业邮箱
QQ 501747125,QQ 3318510886网站建设:合优 网易企业邮箱

 名企招聘
名企招聘
